Modeling the Early Steps of Viral Infection
Viruses have developed efficient molecular tools to hijack the cell transport and transcription machinery. Consequently, they are widely used as gene vectors in many cell types. Yet, they provoke immune responses and synthetic gene vectors are designed but their efficiency is still very low: while one virus can be sufficient to transfer a gene, more than 10,000 synthetic vectors are required. To understand quantitatively the high efficiency of viral particles and their cellular tropism, we develop biophysical models for the early steps of infection. In particular we are interested in modeling the viral escape from the endosome and its cytoplasmic journey until it reaches a nuclear pore.
The Endosomal Escape of Viruses
Proton pumps, that are located on the endosomal membrane, acidify the endosomal compartment and viruses have to escape the endosome before being digested by low-pH activated proteases. On the other hand, new experimental evidences show that a partial denaturation by proteases of the capsid is also required (see Figure 1). Thus, viruses should escape in a certain pH window and quantify the time course of this process is fundamental. Active proteins that change of conformation when they bind a sufficient number of protons, or other low-pH activated ligands such as cathepsins, are responsible of the viral escape. In our biophysical model, we have developed a Markov jump analysis to determine the conformational change rate of these proteins for a given pH. Interestingly, our model nicely fits the data reported for the Influenza hemmagglutinin. Then, we considered small non enveloped viruses, such as parvoviruses, that contain a small number of active proteins in their capsid (about 5) and we assumed the conformational change of one protein should lead to pores formation in the endosomal membrane and disruption of the compartment through an osmotic swelling mechanism. Consequently, neglecting some intermediate steps in the pores formation, we considered the conformational change of one active protein was the limiting event for the escape of all the viruses contained in the endosome and considering a Poissonian entry of protons with a certain rate we computed the escape dynamics of viruses. Interestingly, we found that the mean escape time of Adeno-Associated viruses was around 20 minutes, suggesting they should escape from the late endosomal compartment. In addition, we predict that there should exist an optimal number of viruses in the endosome that maximises the probability they escape in the right pH-range.
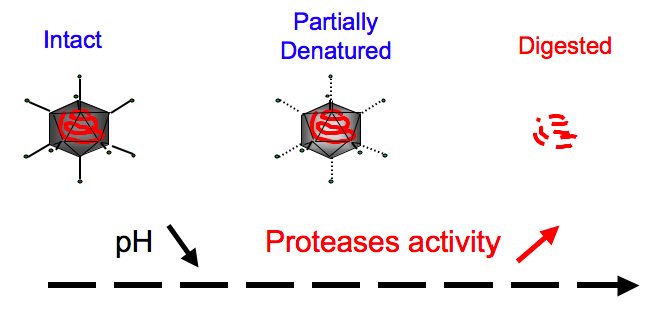
Figure 1: Viruses have to escape in a certain pH-range
Modeling the Cytoplasmic Journey of Viruses
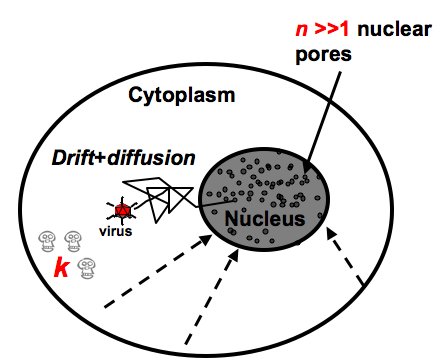 .
. Figure 2: Viral trajectories are coarse-grained in a Langevin stochastic equation that contains both a diffusion and a drift term. Possible degradation or trapping of the viral particle is modeled by a steady state degradation rate k